
Semua kandungan iLive disemak secara perubatan atau fakta diperiksa untuk memastikan ketepatan faktual sebanyak mungkin.
Kami mempunyai garis panduan sumber yang ketat dan hanya memautkan ke tapak media yang bereputasi, institusi penyelidikan akademik dan, apabila mungkin, dikaji semula kajian secara medis. Perhatikan bahawa nombor dalam kurungan ([1], [2], dan lain-lain) boleh diklik pautan ke kajian ini.
Jika anda merasakan bahawa mana-mana kandungan kami tidak tepat, ketinggalan zaman, atau tidak dipersoalkan, sila pilih dan tekan Ctrl + Enter.
Sintesis, rembesan dan metabolisme katekolamin
Pakar perubatan artikel itu

Ulasan terakhir: 06.07.2025
Medula adrenal menghasilkan sebatian dengan struktur yang jauh daripada steroid. Ia mengandungi nukleus 3,4-dihydroxyphenyl (catechol) dan dipanggil katekolamin. Ini termasuk adrenalin, noradrenalin, dan dopamine beta-hydroxytyramine.
Urutan sintesis katekolamin agak mudah: tyrosine → dihydroxyphenylalanine (DOPA) → dopamine → norepinephrine → adrenaline. Tirosin memasuki badan dengan makanan, tetapi juga boleh dibentuk daripada fenilalanin dalam hati di bawah tindakan fenilalanin hidroksilase. Produk akhir penukaran tirosin dalam tisu adalah berbeza. Dalam medulla adrenal, proses itu diteruskan ke peringkat pembentukan adrenalin, di hujung saraf simpatik - norepinephrine, di beberapa neuron sistem saraf pusat, sintesis katekolamin berakhir dengan pembentukan dopamin.
Penukaran tirosin kepada DOPA dimangkinkan oleh tyrosine hydroxylase, yang kofaktornya ialah tetrahydrobiopterin dan oksigen. Adalah dipercayai bahawa enzim ini mengehadkan kadar keseluruhan proses biosintesis katekolamin dan dihalang oleh produk akhir proses tersebut. Tyrosine hydroxylase adalah sasaran utama kesan pengawalseliaan pada biosintesis katekolamin.
Penukaran DOPA kepada dopamin dimangkinkan oleh enzim DOPA decarboxylase (kofaktor - pyridoxal phosphate), yang secara relatifnya tidak spesifik dan menyahkarboksilat asid L-amino aromatik yang lain. Walau bagaimanapun, terdapat tanda-tanda kemungkinan mengubah suai sintesis katekolamin dengan mengubah aktiviti enzim ini. Sesetengah neuron kekurangan enzim untuk penukaran selanjutnya dopamin, dan ia adalah produk akhir. Tisu lain mengandungi dopamine beta-hydroxylase (kofaktor - kuprum, asid askorbik dan oksigen), yang menukar dopamin kepada norepinephrine. Dalam medula adrenal (tetapi tidak di hujung saraf simpatik), phenylethanolamine hadir - metiltransferase yang membentuk adrenalin daripada norepinephrine. Dalam kes ini, S-adenosylmethionine berfungsi sebagai penderma kumpulan metil.
Adalah penting untuk diingat bahawa sintesis phenylethanolamine-N-methyltransferase disebabkan oleh glukokortikoid yang memasuki medula dari korteks melalui sistem vena portal. Ini mungkin menjelaskan fakta bahawa dua kelenjar endokrin yang berbeza digabungkan dalam satu organ. Kepentingan glukokortikoid untuk sintesis adrenalin ditekankan oleh fakta bahawa sel-sel medula adrenal yang menghasilkan noradrenalin terletak di sekitar saluran arteri, manakala sel-sel penghasil adrenalin menerima darah terutamanya dari sinus vena yang terletak di korteks adrenal.
Pecahan katekolamin berlaku terutamanya di bawah pengaruh dua sistem enzim: catechol-O-methyltransferase (COMT) dan monoamine oxidase (MAO). Laluan utama pecahan adrenalin dan noradrenalin secara skematik ditunjukkan dalam Rajah 54. Di bawah pengaruh COMT dengan kehadiran penderma kumpulan metil S-adrenosylmethionine, katekolamin ditukar menjadi normetanephrine dan metanephrine (derivatif 3-O-metil daripada noradrenalin dan adrenalin kemudiannya, yang, di bawah pengaruh MA, di bawah pengaruh MA. kehadiran aldehid oksidase) ke dalam asid vanillylmandelic (VMA), produk pecahan utama noradrenalin dan adrenalin. Dalam kes yang sama, apabila katekolamin pada mulanya terdedah kepada MAO dan bukannya COMT, ia ditukar kepada aldehid 3,4-dioxomandelic, dan kemudian, di bawah pengaruh aldehid oksidase dan COMT, menjadi asid 3,4-dioxomandelic dan VMC. Dengan kehadiran alkohol dehidrogenase, 3-methoxy-4-oxyphenylglycol, yang merupakan produk akhir utama adrenalin dan degradasi noradrenalin dalam CNS, boleh dibentuk daripada katekolamin.
Pecahan dopamin adalah serupa, kecuali metabolitnya kekurangan kumpulan hidroksil pada atom beta-karbon, dan oleh itu, bukannya asid vanillylmandelic, asid homovanillic (HVA) atau asid 3-methoxy-4-hydroxyphenylacetic terbentuk.
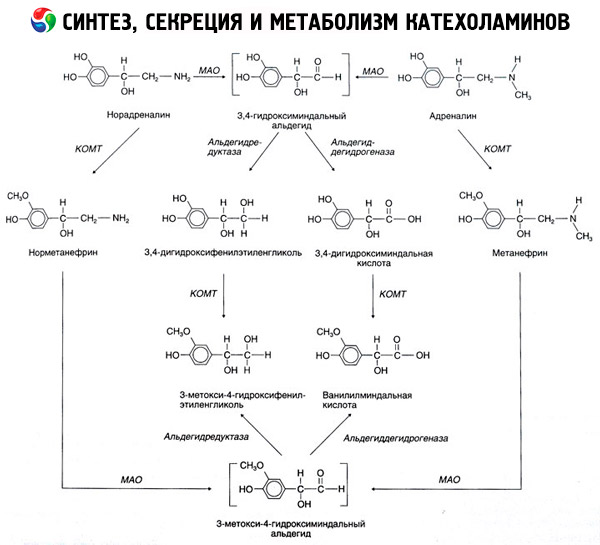
Kewujudan laluan quinoid pengoksidaan molekul katekolamin, yang boleh menghasilkan produk perantaraan dengan aktiviti biologi yang ketara, juga didalilkan.
Norepinephrine dan adrenalin terbentuk di bawah tindakan enzim sitosolik dalam ujung saraf simpatetik dan medula adrenal memasuki butiran rembesan, yang melindungi mereka daripada tindakan enzim degradasi. Penangkapan katekolamin oleh butiran memerlukan perbelanjaan tenaga. Dalam butiran kromafin medulla adrenal, katekolamin terikat rapat dengan ATP (dalam nisbah 4:1) dan protein khusus - kromogranin, yang menghalang penyebaran hormon dari butiran ke dalam sitoplasma.
Rangsangan langsung untuk rembesan katekolamin nampaknya adalah penembusan kalsium ke dalam sel, merangsang eksositosis (penyatuan membran granul dengan permukaan sel dan pecahnya dengan pelepasan lengkap kandungan larut - katekolamin, dopamin beta-hidroksilase, ATP dan kromogranin - ke dalam cecair ekstraselular).
Kesan fisiologi katekolamin dan mekanisme tindakannya
Kesan katekolamin bermula dengan interaksi dengan reseptor tertentu sel sasaran. Walaupun reseptor untuk hormon tiroid dan steroid disetempatkan di dalam sel, reseptor untuk katekolamin (serta hormon asetilkolin dan peptida) terdapat pada permukaan sel luar.
Telah lama diketahui bahawa berhubung dengan beberapa tindak balas, adrenalin atau noradrenalin lebih berkesan daripada isoproterenol katekolamin sintetik, manakala berhubung dengan yang lain, kesan isoproterenol adalah lebih baik daripada tindakan adrenalin atau noradrenalin. Atas dasar ini, konsep telah dibangunkan mengenai kehadiran dua jenis adrenoreceptor dalam tisu: alfa dan beta, dan dalam sesetengahnya hanya satu daripada dua jenis ini boleh hadir. Isoproterenol ialah agonis beta-adrenoreceptor yang paling berkuasa, manakala sebatian sintetik phenylephrine ialah agonis alpha-adrenoreceptors yang paling berkuasa. Katekolamin semulajadi - adrenalin dan noradrenalin - dapat berinteraksi dengan reseptor kedua-dua jenis, tetapi adrenalin mempamerkan pertalian yang lebih besar untuk beta-, dan noradrenalin - untuk reseptor alfa.
Katekolamin mengaktifkan reseptor beta-adrenergik jantung dengan lebih kuat daripada reseptor beta otot licin, yang membenarkan jenis beta dibahagikan kepada subjenis: reseptor beta1 (jantung, sel lemak) dan reseptor beta2 (bronki, saluran darah, dll.). Kesan isoproterenol pada reseptor beta1 melebihi kesan adrenalin dan noradrenalin hanya 10 kali ganda, manakala pada reseptor beta2 ia bertindak 100-1000 kali lebih kuat daripada katekolamin semulajadi.
Penggunaan antagonis khusus (phentolamine dan phenoxybenzamine untuk alpha- dan propranolol untuk beta-reseptor) mengesahkan kecukupan klasifikasi adrenoreceptors. Dopamine mampu berinteraksi dengan kedua-dua reseptor alfa dan beta, tetapi pelbagai tisu (otak, kelenjar pituitari, vesel) juga mempunyai reseptor dopaminergik mereka sendiri, penyekat khususnya adalah haloperidol. Bilangan reseptor beta berbeza dari 1000 hingga 2000 setiap sel. Kesan biologi katekolamin yang dimediasi oleh reseptor beta biasanya dikaitkan dengan pengaktifan adenylate cyclase dan peningkatan kandungan intraselular cAMP. Walaupun reseptor dan enzim disambungkan secara fungsional, ia adalah makromolekul yang berbeza. Guanosine triphosphate (GTP) dan nukleotida purin lain mengambil bahagian dalam modulasi aktiviti adenylate cyclase di bawah pengaruh kompleks reseptor hormon. Dengan meningkatkan aktiviti enzim, ia kelihatan mengurangkan pertalian reseptor beta untuk agonis.
Fenomena peningkatan sensitiviti struktur denervasi telah lama diketahui. Sebaliknya, pendedahan yang berpanjangan kepada agonis mengurangkan sensitiviti tisu sasaran. Kajian tentang reseptor beta telah memungkinkan untuk menerangkan fenomena ini. Telah ditunjukkan bahawa pendedahan yang berpanjangan kepada isoproterenol membawa kepada kehilangan sensitiviti adenilat siklase akibat penurunan bilangan reseptor beta.
Proses penyahpekaan tidak memerlukan pengaktifan sintesis protein dan mungkin disebabkan oleh pembentukan beransur-ansur kompleks reseptor hormon yang tidak dapat dipulihkan. Sebaliknya, pengenalan 6-oxidopamine, yang memusnahkan penghujung simpatik, disertai dengan peningkatan bilangan reseptor beta yang bertindak balas dalam tisu. Ada kemungkinan peningkatan dalam aktiviti saraf simpatik juga menyebabkan desensitisasi berkaitan usia saluran darah dan tisu adipos berhubung dengan katekolamin.
Bilangan adrenoreceptor dalam organ yang berbeza boleh dikawal oleh hormon lain. Oleh itu, estradiol meningkat dan progesteron mengurangkan bilangan alpha-adrenoreceptors dalam rahim, yang disertai dengan peningkatan dan penurunan yang sepadan dalam tindak balas kontraktilnya terhadap katekolamin. Jika "messenger kedua" intraselular yang dibentuk oleh tindakan agonis reseptor beta pastinya cAMP, maka keadaan berkenaan dengan pemancar kesan alfa-adrenergik adalah lebih rumit. Kewujudan pelbagai mekanisme diandaikan: penurunan tahap cAMP, peningkatan kandungan cAMP, modulasi dinamik kalsium selular, dsb.
Untuk menghasilkan semula pelbagai kesan dalam badan, dos adrenalin biasanya diperlukan yang 5-10 kali lebih kecil daripada noradrenalin. Walaupun yang terakhir adalah lebih berkesan berhubung dengan a- dan beta1-adrenoreceptors, adalah penting untuk diingat bahawa kedua-dua katekolamin endogen mampu berinteraksi dengan kedua-dua reseptor alfa dan beta. Oleh itu, tindak balas biologi organ tertentu terhadap pengaktifan adrenergik sebahagian besarnya bergantung pada jenis reseptor yang terdapat di dalamnya. Walau bagaimanapun, ini tidak bermakna pengaktifan selektif pautan saraf atau humoral sistem simpatetik-adrenal adalah mustahil. Dalam kebanyakan kes, peningkatan aktiviti pelbagai pautannya diperhatikan. Oleh itu, secara amnya diterima bahawa hipoglikemia secara refleks mengaktifkan medulla adrenal, manakala penurunan tekanan darah (hipotensi postural) disertai terutamanya oleh pembebasan noradrenalin dari hujung saraf simpatetik.
Adrenoreceptors dan kesan pengaktifannya dalam pelbagai tisu
Sistem, organ |
Jenis reseptor adrenergik |
Reaksi |
Sistem kardiovaskular: |
||
Hati |
Beta |
Peningkatan kadar denyutan jantung, kekonduksian dan pengecutan |
Arteriol: |
||
Kulit dan membran mukus |
Alfa |
Pengurangan |
Otot rangka |
Beta |
Pengecutan Pengembangan |
Organ perut |
Alpha (lagi) |
Pengurangan |
Beta |
Sambungan |
|
Urat |
Alfa |
Pengurangan |
Sistem pernafasan: |
||
Otot bronkus |
Beta |
Sambungan |
Sistem penghadaman: |
||
Perut |
Beta |
Kemahiran motor berkurangan |
Usus |
Alfa |
Penguncupan sfinkter |
Limpa |
Alfa |
Pengurangan |
Beta |
Relaksasi |
|
Pankreas eksokrin |
Alfa |
Rembesan berkurangan |
Sistem urogenital: |
Alfa |
Penguncupan sphincter |
Pundi kencing |
Beta |
Kelonggaran otot ejektor |
Alat kelamin lelaki |
Alfa |
Ejakulasi |
Mata |
Alfa |
Pelebaran murid |
Kulit |
Alfa |
Berpeluh bertambah |
Kelenjar air liur |
Alfa |
Perkumuhan kalium dan air |
Beta |
Rembesan amilase |
|
Kelenjar endokrin: |
||
Pulau kecil pankreas |
||
Sel beta |
Alpha (lagi) |
Rembesan insulin berkurangan |
Beta |
Peningkatan rembesan insulin |
|
Sel alfa |
Beta |
Peningkatan rembesan glukagon |
8-sel |
Beta |
Peningkatan rembesan somatostatin |
Hipotalamus dan kelenjar pituitari: |
||
Somatotrof |
Alfa |
Peningkatan rembesan STH |
Beta |
Pengurangan rembesan STH |
|
Laktotrof |
Alfa |
Pengurangan rembesan prolaktin |
Thyrotrophs |
Alfa |
Rembesan TSH berkurangan |
Kortikotrof |
Alfa |
Peningkatan rembesan ACTH |
| beta | Rembesan ACTH berkurangan | |
Kelenjar tiroid: |
||
Sel folikel |
Alfa |
Pengurangan rembesan tiroksin |
Beta |
Peningkatan rembesan tiroksin |
|
Sel parafolikular (K). |
Beta |
Peningkatan rembesan calcitonin |
Kelenjar paratiroid |
Beta |
Peningkatan rembesan PTH |
Buah pinggang |
Beta |
Peningkatan rembesan renin |
Perut |
Beta |
Peningkatan rembesan gastrin |
BX |
Beta |
Peningkatan penggunaan oksigen |
Hati |
? |
Peningkatan glikogenolisis dan glukoneogenesis dengan pembebasan glukosa; peningkatan ketogenesis dengan pembebasan badan keton |
Tisu adiposa |
Beta |
Peningkatan lipolisis dengan pembebasan asid lemak bebas dan gliserol |
Otot rangka |
Beta |
Peningkatan glikolisis dengan pembebasan piruvat dan laktat; menurunkan proteolisis dengan penurunan dalam pelepasan alanin, glutamin |
Adalah penting untuk mengambil kira bahawa keputusan pentadbiran intravena katekolamin tidak selalu mencerminkan kesan sebatian endogen dengan secukupnya. Ini terpakai terutamanya kepada norepinephrine, kerana dalam badan ia dilepaskan terutamanya bukan ke dalam darah, tetapi terus ke celah sinaptik. Oleh itu, norepinephrine endogen mengaktifkan, sebagai contoh, bukan sahaja reseptor alfa vaskular (peningkatan tekanan darah), tetapi juga reseptor beta jantung (peningkatan kadar denyutan jantung), manakala pengenalan norepinephrine dari luar membawa terutamanya kepada pengaktifan reseptor alfa vaskular dan refleks (melalui vagus) memperlahankan degupan jantung.
Dos adrenalin yang rendah mengaktifkan terutamanya reseptor beta saluran otot dan jantung, mengakibatkan penurunan rintangan vaskular periferi dan peningkatan output jantung. Dalam sesetengah kes, kesan pertama mungkin berlaku, dan hipotensi berkembang selepas pentadbiran adrenalin. Dalam dos yang lebih tinggi, adrenalin juga mengaktifkan reseptor alfa, yang disertai dengan peningkatan rintangan vaskular periferal dan, dengan latar belakang peningkatan output jantung, membawa kepada peningkatan tekanan arteri. Walau bagaimanapun, kesannya terhadap reseptor beta vaskular juga dipelihara. Akibatnya, peningkatan tekanan sistolik melebihi penunjuk tekanan diastolik yang sama (peningkatan tekanan nadi). Dengan pengenalan dos yang lebih tinggi, kesan alpha-mimetik adrenalin mula diguna pakai: peningkatan tekanan sistolik dan diastolik secara selari, seperti di bawah pengaruh norepinephrine.
Kesan katekolamin pada metabolisme terdiri daripada kesan langsung dan tidak langsung. Yang pertama direalisasikan terutamanya melalui reseptor beta. Proses yang lebih kompleks dikaitkan dengan hati. Walaupun peningkatan glikogenolisis hati secara tradisinya dianggap sebagai hasil daripada pengaktifan reseptor beta, terdapat juga bukti penglibatan reseptor alfa. Kesan tidak langsung katekolamin dikaitkan dengan modulasi rembesan banyak hormon lain, seperti insulin. Dalam kesan adrenalin pada rembesannya, komponen alfa adrenergik jelas mendominasi, kerana telah ditunjukkan bahawa sebarang tekanan disertai dengan perencatan rembesan insulin.
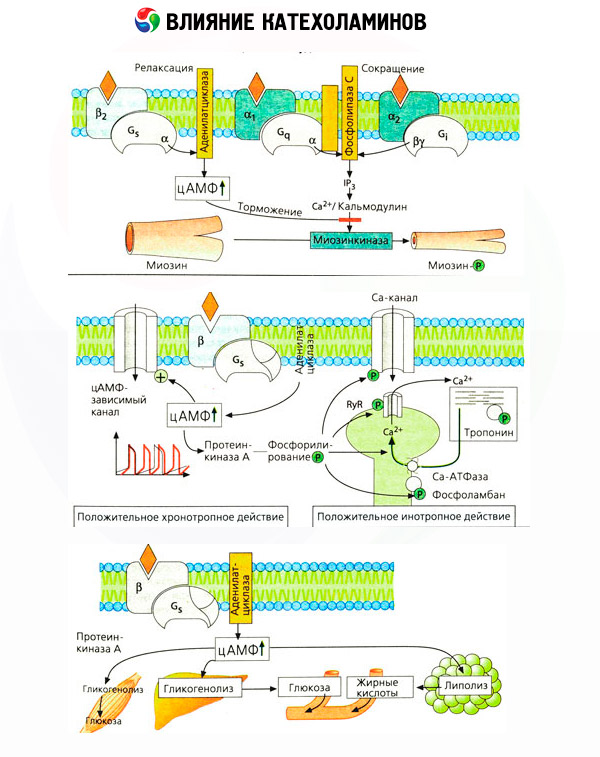
Gabungan kesan langsung dan tidak langsung katekolamin menyebabkan hiperglikemia, yang dikaitkan bukan sahaja dengan peningkatan pengeluaran glukosa hepatik, tetapi juga dengan perencatan penggunaannya oleh tisu periferi. Pecutan lipolisis menyebabkan hiperlipasidemia dengan peningkatan penghantaran asid lemak ke hati dan peningkatan pengeluaran badan keton. Peningkatan glikolisis dalam otot membawa kepada peningkatan dalam pelepasan laktat dan piruvat ke dalam darah, yang, bersama-sama dengan gliserol yang dikeluarkan dari tisu adipos, berfungsi sebagai prekursor glukoneogenesis hepatik.
Peraturan rembesan katekolamin. Persamaan produk dan kaedah tindak balas sistem saraf simpatetik dan medula adrenal adalah asas untuk menggabungkan struktur ini menjadi satu sistem simpatetik-adrenal badan dengan peruntukan pautan saraf dan hormonnya. Pelbagai isyarat aferen tertumpu di hipotalamus dan pusat saraf tunjang dan medulla oblongata, dari mana mesej eferen berasal, beralih ke badan selular neuron preganglionik yang terletak di tanduk sisi saraf tunjang pada tahap serviks VIII - segmen lumbar II-III.
Akson preganglionik sel-sel ini meninggalkan saraf tunjang dan membentuk sambungan sinaptik dengan neuron yang terletak di ganglia rantai simpatetik atau dengan sel-sel medula adrenal. Gentian preganglionik ini adalah kolinergik. Perbezaan asas pertama antara neuron postganglionik bersimpati dan sel kromafin medulla adrenal ialah yang kedua menghantar isyarat kolinergik yang diterima oleh mereka bukan melalui pengaliran saraf (saraf adrenergik postganglionik), tetapi melalui laluan humoral, melepaskan sebatian adrenergik ke dalam darah. Perbezaan kedua ialah saraf postganglionik menghasilkan norepinephrine, manakala sel-sel medulla adrenal menghasilkan terutamanya adrenalin. Kedua-dua bahan ini mempunyai kesan yang berbeza pada tisu.